Цепь переноса водорода и электрона (дыхательная цепь). Комплексы переноса
электронов. Окислительное фосфорилирование. Хемиосмотическая теория окисления и
фосфорилирования. Механизмы сопряжения процеСтраница 1

Цикл Кребса, глиоксилатный и пентозофосфатный пути функционируют только в условиях достаточного количества кислорода. В то же время 02 непосредственно не участвует в реакциях этих циклов. Точно так же в перечисленных циклах не синтезируется АТФ (за исключением АТФ, образующегося в цикле Кребса в результате субстратного фосфорилирования на уровне сукцинил-СоА).
Кислород необходим для заключительного этапа дыхательного процесса, связанного с окислением восстановленных коферментов НАДH и ФАДH2 в дыхательной электронтранспортной цепи (ЭТЦ) митохондрий. С переносом электронов по ЭТЦ сопряжен и синтез АТФ.
Дыхательная ЭТЦ, локализованная во внутренней мембране митохондрий, служит для передачи электронов от восстановленных субстратов на кислород, что сопровождается трансмембранным переносом ионов Н + . Таким образом, ЭТЦ митохондрий (как и тилакоидов) выполняет функцию окислительно-восстановительнои Н -помпы. ,
Б. Чане и др. (США) в 50-х годах, используя значения окислительно-восстановительных потенциалов известных в то время переносчиков е-, спектрофотометрические данные о временной последовательности их восстановления и результаты ингибиторного анализа, расположили компоненты ЭТЦ митохондрий в следующем порядке:
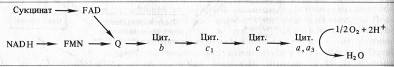
Пара электронов от НАДH или сукцината передается по ЭТЦ до кислорода, который, восстанавливаясь и присоединяя два протона, образует воду.
Д. Грин (1961) пришел к выводу, что все переносчики электронов в митохондриальной мембране сгруппированы в четыре комплексу, что было подтверждёно дальнейшими исследованиями.
Согласно современным данным дыхательная цепь митохондрий включает в себя четыре основных мультиэнзимных комплекса и два небольших по молекулярной массе компонента — убихинон и цитохром с
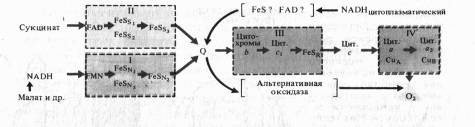
Рис.7. Дыхательная электронтранспортная цепь растительных митохондрий
Комплекс I осуществляет перенос электронов от НАДH к убихинону Q. Его субстратом служат молекулы вдутримитохондриального НАДH, восстанавливающиеся в цикле Кребса. Всостав комплекса входит флавиновая ФМН-зависимая НАДH: убихи- нон-оксидоредуктаза, содержащая три железосерных центра (FeSN1-3). При встраивании в искусственную фосфолипидную мембрану этот комплекс функционирует как протонная помпа.
Комплекс II катализирует окисление сукцината убихиноном. Эту функцию осуществляют флавиновая (ФАД-зависимая) сукцинат : убихинон-оксидоредуктаза, в состав которой также входят три железосерных центра (Fes1_3).
Koмплекс III переносит электроны от восстановленного убихинона к цитохрому с, т. е. функционирует как убихинол: цитохром Т-оксидредуктаза. В своем составе он содержит цитохромы
b556
и
b560
,
цитохром с, и железосерный белок Риске. По структуре и функции этот комплекс сходен с цитохромным комплексом
b6
—
f тилакоидов хлоропластов. В присутствии убихинона комплекс III осуществляет активный трансмембранный перенос протонов.
В терминальном комплексе IV электроны переносятся от цитохрома
с к кислороду. _т. е. этот комплекс является цитохромом с: кислород-оксидоредуктазой (цитохромоксидазой). В его состав входят четыре редокс-компонента: цитохромы а
и а3
и два атома меди. Цитохром а3 и Сив способны взаимодействовать с 02, на который передаются электроны с цитохрома а — СиА. Транспорт электронов через комплекс IV сопряжен с активным транспортом ионов Н +.
В последние годы в результате изучения пространственного расположения компонентов ЭТЦ во внутренней мембране митохондрии показано что комплексы I, III и IV пересекают мембрану. На внутренней стороне мембраны, обращенной к матриксу, два электрона и два протона от НАДH поступают на флавинмононуклеотид комплекса I.
Электроны передаются на FeS-центры. Пара электронов от FeS-центров захватывается двумя молекулами окисленного убихинона, которые принимают два иона Н + , образуя семихиноны (2QH) и диффундируя к комплексу III. На эти семихиноны поступает еще пара электронов от цитохрома Ь560 комплекса III, что делает возможной реакцию семихинонов с еще двумя протонами из мАТФикса с образованием 2QH2. Полностью восстановленный убихинон (убихинол) отдает 2е~ цитохрому Ь556 и 2е~ FeSR — цитохрому с,. В результате освобождаются четыре иона Н+, выходящие в межмембранное пространство митохондрии. Окисленные молекулы убихинона вновь диффундируют к комплексу I и готовы принимать от него (или от комплекса II) новые электроны и протоны. Таким образом, цитохромы Ь служат донорами двух электронов для переноса двух дополнительных протонов через липидную фазу мембраны на каждые два электрона, поступающие из комплекса I.
Использование интеркалирующих агентов.
Этот способ детекции основан на том факте, что флуоресценция бромистого этидия и SYBR Green I значительно возрастает при их внедрении в двухцепочечные молекулы ДНК [14] (Рис. 4). Таким образом, можно наблюдать за накоплением продуктов амплификации. Очень важно отметить то, что увеличение флуоресценции может быть связано как с накопление ...
Великие массовые вымирания
Предполагается, что первыми зафиксированными событиями такого типа были вымирания вендского периода. На рубеже рифея и венда отмечается резкое обеднение фитопланктона, что было связано с влиянием одного из величайших в истории Земли оледенений. После оледенения появились разнообразные крупные многоклеточные, которые очень быстро распрос ...
Вымокание
Вымокание проявляется преимущественно весной в пониженных местах в период таяния снега, реже во время длительных оттепелей, когда на поверхности почвы накапливается талая вода, которая не впитывается в замершую почву и может затопить растения. В этом случае причиной гибели растений служит резкий недостаток кислорода (анаэробные условия ...