Цепь переноса водорода и электрона (дыхательная цепь). Комплексы переноса
электронов. Окислительное фосфорилирование. Хемиосмотическая теория окисления и
фосфорилирования. Механизмы сопряжения процеСтраница 2

Водорастворимый цитохром с на наружной стороне мембраны, получив 2е~ от FeSR — цитохрома ciy передает их на цитохром а — Сид комплекса IV. Цитохром а3 — Сив, связывая кислород, переносит на него эти электроны, в результате чего с участием двух протонов образуется вода. Как уже отмечалось, цитохромоксидазный комплекс способен также переносить ионы Н+ через митохондриальную мембрану.
Таким образом, из матрикса митохондрии при транспорте каждой пары электронов от НАДH к 1/2 02 в трех участках ЭТЦ (комплексы I, III, IV) через мембрану наружу переносятся по крайней мере шесть протонов. Как будет показано далее, именно в этих трех участках окислительные процессы в ЭТЦ сопрягаются с синтезом АТФ. Передача 2е~ от сукцината на убихинон в комплексе II не сопровождается трансмембранным переносом протонов. Это приводит к тому, что при использовании сукцината в качестве дыхательного субстрата в ЭТЦ остаются лишь два участка, в которых функционирует протонная помпа.
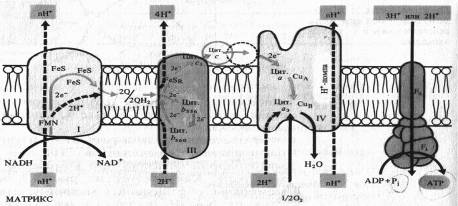
Рис. 8. Предполагаемое расположение компонентов цепи переноса электронов во внутренней мембране митохондрий в соответствии с хемиосинтетической теорией Митчелла
Особенностью растительных митохондрий (отличающей их от митохондрий животных) является способность окислять экзогенный НАДH, т. е. НАДH, поступающий из цитоплазмы. Это окисление осуществляется по крайней мере двумя флавиновыми НАДH-дегидрогеназами, из которых одна локализована на наружной стороне внутренней мембраны митохондрий, а другая — в их наружной мембране. Первая из них передает электроны в ЭТЦ митохондрий на убихинон, а вторая — на цитохром с. Для функционирования НАДH- дегидрогенэзы на наружной стороне внутренней мембраны необходимо присутствие кальция.
Другое существенное отличие растительных митохондрий состоит в том, что во внутренней мембране помимо основного (цитохромного) пути переноса электронов имеется альтернативный путь переноса е~, устойчивый к действию цианида.
Перенос электронов от НАДH к молекулярному кислороду через ЭТЦ митохондрий сопровождается потерей свободной энергии. Какова судьба этой энергии? Еще в 1931 г. В. А. Энгельгардт показал, что при аэробном дыхании накапливается АТФ. Он первый высказал идею о сопряжении между фосфорилированием АДФ и аэробным дыханием. В 1937—1939 гг. биохимики В. А. Белицер в СССР и Г. Калькар в США установили, что при окислении промежуточных продуктов цикла Кребса, в частности янтарной и лимонной кислот, суспензиями животных тканей исчезает неорганический фосфат и образуется АТФ. В анаэробных условиях или при подавлении дыхания цианидом такого фосфорилирования не происходит. Процесс фосфорилирования АДФ с образованием АТФ, сопряженный с переносом электронов по ЭТЦ митохондрий, получил название окислительного фосфорилирования
.
По поводу механизма окислительного фосфорилирования существуют три теории: химическая, механохимическая (конформационная) и хемиосмотическая.
Химическая и механохимическая гипотезы сопряжения.
Согласно химической гипотезе в митохондриях имеются интермедиаторы белковой природы (X, Y, Z), образующие комплексы с соответствующими восстановленными переносчиками. В результате окисления переносчика в комплексе возникает высокоэнергетическая связь. При распаде комплекса к интермедиатору с высокоэнергетической связью присоединяется неорганический фосфат, который затем передается на АДФ:
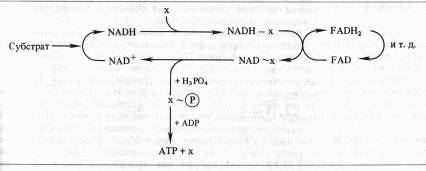
Однако несмотря на упорные поиски, не удалось выделить или как-то иначе доказать реальное существование постулированных высокоэнергетических интермедиаторов типа X ~ P. Гипотеза химического сопряжения не объясняет, почему окислительное фосфорилирование обнаруживается только в препаратах митохондрий с ненарушенными мембранами. И, наконец, с позиций этой гипотезы не находит объяснения способность митохондрий подкислять внешнюю среду и изменять свой объем в зависимости от степени их энергизации.
Способность митохондриальных мембран к конформационным изменениям и связь этих изменений со степенью энергизации митохондрий послужила основой для создания механохимических гипотез образования АТФ в ходе окислительного фосфорилирования. Согласно этим гипотезам энергия, высвобождающаяся в процессе переноса электронов, непосредственно используется для перевода белков внутренней мембраны митохондрий в новое, богатое энергией конформационное состояние, приводящее к образованию АТФ. Одна из гипотез подобного рода, выдвинутая американским биохимиком П. Д. Бойером (1965), может быть представлена в виде следующей схемы:
Великие массовые вымирания
Предполагается, что первыми зафиксированными событиями такого типа были вымирания вендского периода. На рубеже рифея и венда отмечается резкое обеднение фитопланктона, что было связано с влиянием одного из величайших в истории Земли оледенений. После оледенения появились разнообразные крупные многоклеточные, которые очень быстро распрос ...
Прокариоты
Прокариоты – это простейшие одноклеточные организмы, которым для того, чтобы выжить, требуется лишь благоприятная химическая среда.
Некоторые необходимые понятия.
Репрессор – это белок, блокирующий транскрипцию гена. В lac-системе репрессор представляет собой тетрамерный белок, и называется lac-репрессором. Он связывается с определенн ...
Материалы исследования
Объектом исследования служила капиллярная кровь, которая забиралась в гепаринизированный капилляр в объеме 200 мкл. В полученных образцах определяли рН, рСО2, уровень НСО3-, ВЕ, концентрацию лактата.
Экспериментальную группу составили пловцы с квалификацией: мастера спорта и мастера спорта международного класса. Спортсмены выполняли те ...