Цепь переноса водорода и электрона (дыхательная цепь). Комплексы переноса
электронов. Окислительное фосфорилирование. Хемиосмотическая теория окисления и
фосфорилирования. Механизмы сопряжения процеСтраница 3

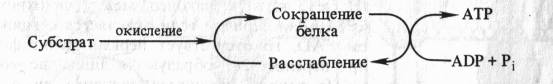
Автор предположил, что запасание энергии происходит путем конформационных изменений ферментов ЭТЦ аналогично тому, как это наблюдается в белках мышц. Актомиозиновый комплекс сокращается, гидролизуя АТФ. Если сокращение белкового комплекса достигается за счет другой формы энергии (за счет окисления), то расслабление, возможно, будет сопровождаться синтезом АТФ.
Таким образом, согласно механохимическим гипотезам, энергия окисления превращается сначала в механическую энергию, а затем в энергию высокоэнергетической связи АТФ. Однако, подобно химической теории сопряжения, механохимические гипотезы также не могут объяснить подкисление митохондриями окружающей среды.
Хемиосмотическая теория сопряжения.
В настоящее время наибольшим признанием пользуется хемиосмотическая теория английского биохимика П. Митчелла (1961). Он высказал предположение, что поток электронов через систему молекул-переносчиков сопровождается транспортом ионов Н+ через внутреннюю мембрану митохондрий. В результате на мембране создается электрохимический потенциал ионов Н + , включающий химический, или осмотический, градиент и электрический градиент (мембранный потенциал). Согласно хемиосмотической теории электрохимический трансмембранный потенциал ионов Н+ и является источником энергии для синтеза АТФ за счет обращения транспорта ионов Н+ через протонный канал мембранной Н + -АТФазы.
Теория Митчелла исходит из того, что переносчики перешнуровывают мембрану, чередуясь таким образом, что в од сторону возможен перенос и электронов, и протонов, а в об ратную — только электронов. В результате ионы Н+ накапливаются на одной стороне мембраны.
Между двумя сторонами внутренней митохондриальной мембраны в результате направленного движения протонов против концентрационного градиента возникает электрохимический потенциала Энергия, запасенная таким образом, используется для синтеза АТФ как результат разрядки мембраны при обратном (по концентрационному градиенту) транспорте протонов через АТФазу, которая работает в этом случае как АТФ-синтетаза.
За прошедший период хемиосмотическая гипотеза Митчелла получила целый ряд экспериментальных подтверждений. Одним из доказательств роли протонного градиента в образовании АТФ при окислительном фосфорилировании может служить разобщающее действие на этот процесс некоторых веществ. Известно, что 2,4-динитрофенол (2,4-ДНФ) подавляет синтез АТФ, но стимулирует транспорт электронов (поглощение 02), т. е. разобщает дыхание (окисление) и фосфорилирование. Митчелл предположил, что такое действие 2,4-ДНФ связано с тем, что он переносит протоны через мембрану (т. е. является протонофором) и поэтому разряжает ее. Это предположение полностью подтвердилось. Оказалось, что разные по своей химической природе вещества, разобщающие окисление и фосфорилирование, сходны в том, что, во-первых, они растворимы в липидной фазе мембраны, а, во-вторых, это слабые кислоты, т. е. легко приобретают и теряют протон в зависимости от рН среды. В. П. Скулачев на искусственных фосфолипидных мембранах показал, что чем легче вещество переносит протоны через мембрану, тем сильнее разобщает эти процессы. Другое экспериментальное подтверждение роли протонного градиента в фосфорилировании было получено Митчеллом, который сообщил о синтезе АТФ в митохондриях в результате замены щелочной инкубационной среды на кислую (т. е. в условиях искусственно созданного трансмембранного градиента ионов Н+).
В 1973 г. Э. Рэкеру (США) удалось получить липосомы (везикулы из фосфолипидов), в которые была встроена АТФаза, выделенная из митохондрий сердца быка, и хромопротеин галофильной бактерии Halobacterium halobium — бактериородопсин, обусловливающий создание протонного градиента за счет энергии света. Фосфолипиды для реконструкции мембран этих липосом были выделены из растений (соевые бобы). Полученные таким образом гибридные пузырьки на свету осуществляли фосфорилирование.
Ферменты сыворотки крови
В основе многих патологических и предпатологических состояний организма лежат нарушения функционирования ферментных систем. Многие ферменты локализуются внутри клеток, а поэтому в сыворотке (плазме) крови их активность низка или вообще отсутствует. Именно поэтому анализируя внеклеточные жидкости (кровь), по активности определенных ферме ...
Химический состав
По химическому составу бактерии не отличаются от клеток других организмов. Бактериальная клетка содержит 80% воды и 20% сухого остатка. Около 90% сухого остатка бактерии составляют высокомолекулярные соединения: нуклеиновые кислоты (10%), белки (40%), полисахариды (15%}, пептидогликон (10%) и липиды (15%); остальные 10% приходятся на мо ...
Скелет
Череп рыб неподвижно соединен с позвоночником, образуя единый клин, легко входящий в толщу воды. Позвоночник имеет только два отдела: туловищный и хвостовой. Двояковогнутые позвонки соединяются при помощи суставных отростков, обеспечивая прочность осевого скелета и его подвижность.
Череп подвижно соединен с позвоночником. Позвоночник д ...