Ответы сложных клеток

При отведении сигналов от отдельных нейронов зрительной коры кроме простых клеток также можно обнаружить и клетки, которые отличаются по своему поведению. Эти сложные клетки, которые преобладают в слоях 2, 3 и 5, имеют два важных свойства, общих с простыми клетками: освещение всего их рецептивного поля неэффективно, и им необходима определенная ориентация границы света и тени в пределах поля. Однако они не так критичны к точному расположению стимула, как простые клетки. Более того, у них также нет четких "on"- и "off"-зон, которые можно определить с помощью световых пятен. Большинство сложных клеток отвечает импульсацией на протяжении всего времени, пока определенным образом ориентированный пучок света падает в пределах границ рецептивного поля, как показано в примере на рис. 4 Значение же сигнала, поступающего от сложных клеток, значительно отличается от сигналов от простых клеток. Простая клетка локализует определенным образом ориентированную полосу света, расположенную в определенной области рецептивного поля; сложная клетка посылает сигналы в ответ на определенную ориентацию, абстрагируясь от четкой локализации стимула в пределах рецептивного поля.
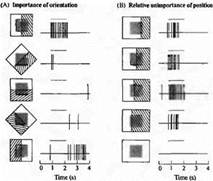
Рис. 4 Ответы сложных клеток в полосатой коре кошки. Клетка реагирует наилучшим образом на вертикальную границу, проходящую через ее рецептивное поле (серый квадрат в центре). (А) При освещении левой половины и затемнении (заштриховано) справа (первая запись) возникает "on"-ответ. При освещении справа (пятая запись) возникает "off"-ответ. Любая ориентация освещения, отличная от вертикальной, менее эффективна. (В) Расположение границы света в пределах поля не играет существенной роли. Освещение всего рецептивного поля (самая нижняя запись) не вызывает никакого ответа.
Можно различить два основных класса сложных клеток, причем оба они наилучшим образом реагируют на перемещение краев или узких полосок света фиксированной ширины и определенной ориентации. Один тип клеток отвечает сигналами, показанными на рис. 4 Сигналы возникают в ответ на границу света определенной ориентации в любом месте в пределах ограниченной области. В этом примере вертикальная граница света вызывает практически одинаковые по амплитуде сигналы в любой из четырех локализаций, как показано на рис. 4 Другие ориентации границы являются неэффективными. Для клеток этого типа ответы становятся более стабильными и высокоамплитудными при удлинении размеров границы или узкого пучка до определенного предела, после которого параметры сигналов уже не меняются.
Другие сложные клетки, подобно конечно-прерывистым простым клеткам (end-stopped simple cells), требуют узких пучков или краев, которые кончаются в пределах их рецептивного поля. Наилучшим стимулом для таких клеток, следовательно, будет не только определенная ориентация, но и какая-то прерывистость, такая как прерывание линии, внутренний или наружный угол (angle and corner) какого-либо контура. Однако диффузное освещение правого поля не дает ослабление сигнала (последний сигнал на рисунке). Таким образом, это нельзя назвать "off"-зоной. Оптимальным стимулом для этой зоны является наружный угол. Более того, стимул должен перемещаться в определенную сторону. Такая чувствительность к направлению перемещения — свойство, как правило, присущее сложным клеткам. Однако можно найти и более требовательные сложные клетки, особенно в пределах экстрастриарной коры).
Асиалогликопротеиновый рецептор
Этот рецептор группы I, называемый также печеночным лектином, связывается с десиалированными гликопротеинами сыворотки и узнает гликопротеины с галактозой или N‑ацетилгалактозой на конце. Следует отметить две интересные особенности: 1) рецептор имеет единственный трансмембранный сегмент и является одним из нескольких рецепторов, у ...
Основы теории самоорганизации систем
Состояние системы зависит от ее параметров и множества внутренних и внешних факторов. Например, для нахождения возможных вариантов колебаний физического маятника нужно знать всего два параметра – координату и скорость. Их значение в любой момент времени будет определятся свойствами самого маятника (длина его подвеса, масса и т.д.) и вне ...
Предложите гипотезу о причинах возникновения в ходе эволюции лишайников
Одной из гипотез о причинах возникновения лишайников в процессе эволюции может быть гипотеза о постепенном переходе от паразитизма к симбиозу. Основной компонент лишайников - гриб - формирует гаустории, которыми гриб внедряется в клетки водоросли и поглощает их содержимое. Вероятно, на первом этапе возникновения лишайников такой паразит ...