АТФ как основная энергетическая валюта клетки, её структура и функции. Механизмы
синтеза АТФСтраница 1

Процессы обмена вещества включают в себя реакции, идущие с потреблением энергии, и реакции с выделением энергии. В некоторых случаях эти реакции сопряжены. Однако часто реакции, в которых энергия выделяется, отделены в пространстве и во времени от реакций, в которых она потребляется. В процессе эволюции у растительных и животных организмов выработалась возможность хранения энергии в форме соединений, обладающих богатыми энергией-связями. Среди них центральное место занимает аденозинтрифосфат (АТФ). АТФ представляет собой нуклеотидфосфат, состоящий из азотистого основания (аденина), пентозы (рибозы) и трех молекул фосфорной кислоты. Две концевые молекулы фосфорной кислоты образуют макроэргические, богатые энергией связи. В клетке АТФ содержится главным образом в виде комплекса с ионами магния. Аденозинтрифосфат в процессе дыхания образуется из аденозиндифосфата и остатка неорганической фосфорной кислоты (Фн) с использованием энергии, освобождающейся при окислении различных органических веществ:
АДФ + ФН --> АТФ + Н2О
При этом энергия окисления органических соединении превращается в энергию фосфорной связи.
В 1939—1940 гг. Ф. Липман установил, что АТФ служит главным нереносчиком энергии в клетке. Особые свойства этого вещества определяются тем, что конечная фосфатная группа легко переноситься с АТФ на другие соединения или отщепляется с выделением энергии, которая может быть использована на физиологические функции. Эта энергия представляет собой разность между свободной энергией АТФ и свободной энергией образующихся продуктов (AG). AG — это изменение свободной энергии системы или количество избыточной энергии, которая освобождается при реорганизации химических связей. Распад АТФ происходит по уравнению AТФ + Н20 = АДФ + ФН, при этом происходит как бы разрядка аккумулятора, при рН 7 выделяется AG = —30,6 кДж. Этот процесс катализируется ферментом аденозинтрифосфатазой - (АТФ-аза) Равновесие гидролиза АТФ смещено в сторону завершения peaкции, что и обусловливает большую отрицательную величину свободной энергии гидролиза. Это связано с тем, что при диссоциации. Четырех гидроксильных группировок при рН 7 АТФ имеет четыре отрицательных заряда. Близкое расположение зарядов друг к другу способствует их отталкиванию и, следовательно, отщеплению фосфатных группировок. В результате гидролиза образуются соединения с одноименным зарядом (АДФ3~ и НР04~), которые отживаются друг от друга, что препятствует их соединению. Уникальные свойства АТФ объясняются не только тем, что при ее гидролизе выделяется большое количество энергии, но и тем, что она обладает способностью отдавать концевую фосфатную группу вместе с запасом энергии на другие органические соединения. Энергия, заключенная в макроэргической фосфорной связи, используется на физиологическую деятельность клетки. Вместе с тем по величине свободной энергии гидролиза — 30,6 кДж/моль АТФ занимает промежуточное положение. Благодаря этому система АТФ — АДФ может служить носчиком фосфатных групп от фосфорных соединений с более высокой энергией гидролиза, например фосфоенолпируват (53,6 К/моль), к соединениям с более низкой энергией гидролиза, пример сахарофосфатам (13,8 кДж/моль). Таким образом, система АДФ является как бы промежуточной или сопрягающей.
Механизм синтеза АТФ. Сопряжение диффузии протонов назад через внутреннюю мембрану митохондрии с синтезом АТФ осуществляется с помощью АТФазного комплекса, получившего название фактора сопряжения F,. На электронно- микроскопических снимках эти факторы выглядят глобулярными образованиями грибовидной формы на внутренней мембране митохондрий, причем их «головки» выступают в матрикс. F1 — водорастворимый белок, состоящий из 9 субъединиц пяти различных типов. Белок представляет собой АТФазу и связан с мембраной через другой белковый комплекс F0, который перешнуровывает мембрану. F0 не проявляет каталитической активности, а служит каналом для транспорта ионов Н+ через мембрану к Fx.
Механизм синтеза АТФ в комплексе Fi~ F0 до конца не выяснен. На этот счет имеется ряд гипотез.
Одна из гипотез, объясняющих образование АТФ посредством так называемого прямого механизма, была предложена Митчеллом.
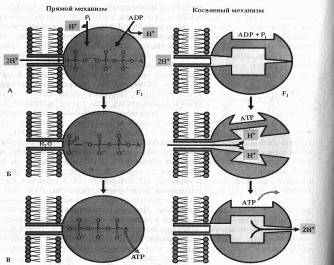
Рис. 9. Возможные механизмы образования АТФ в комплексе F1 – F0
Субстраты для синтеза эйкозаноидов
Главный субстрат для синтеза эйкозаноидов у человека - арахидоновая кислота, так как её содержание в организме человека значительно больше остальных полиеновых кислот - предшественников эйкозаноидов.
В образовании эйкозанондов принимают участие также и другие незаменимые жирные кислоты (линолевая и а-линоленовая), но только после элонг ...
Выявление противогрибковых AT и грибковых Аг
Наиболее часто применяют реакцию латекс-агглютинации (выявляет IgM), PCK (выявляет IgG) и ИФА. Результаты реакций часто могут быть сомнительными вследствие перекрёстного реагирования с Аг различных грибов. Тем не менее идентификация AT или циркулирующих Аг в крови, СМЖ и моче позволяет установить диагноз до получения результатов посевов ...
Биометрическая обработка больших выборок (X+- mx, Cv, t)
Биометрия – наука о способах применения математическ, статистическ методов в биологии. Выборка – часть генеральной совокупности, кот исследуется с целью характеристики всего массива. (Cv
= σ / x), (t
= x/m), (x
= A+b·l), (b
=(∑p·a)/n), (m
= σ/√n), (x+-2,5·σ), σ
=l·√((∑p·a²)/n) - b²) ...