Стратегии изучения коры

Каким образом сигналы, определяющие маленькие, яркие, темные или цветовые пятна на сетчатке могут быть преобразованы в сигналы, которые несут информацию о форме, размере, цвете, движении и глубине расположения объектов. Методики, считающиеся сейчас рутинными, такие как оптическая регистрация, введение пероксидазы хрена или сканирование мозга, в ту пору даже не могли быть представлены. В начале работы перед Хьюбелем и Визелем стояли совершенно неизученные вопросы, которые исходили из их предположения, что зрительные центры в коре осуществляют переработку информации подобно тому, как это происходит в сетчатке, только на более высоком уровне.
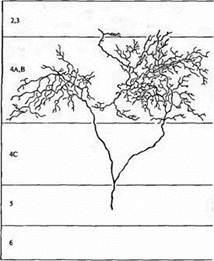
Рис. 1. Аксон "off" центра из ЛКТ, заканчивающийся в слое 4 зрительной коры кошки. В аксон при помощи микроэлектрода была введена пероксидаза хрена. Терминали сгруппированы в два кластера, разделенных неокрашенной зоной, получающей иннервацию от другого глаза.
Одной из ключевых стратегий в их анализе было использование стимулов, которые имитировали сигналы, возникающие в нормальных условиях. Например, края, контуры, а также простые паттерны, представленные глазу, позволяли выявлять особенности организации, которые никогда бы не были обнаружены, используя яркие вспышки света, не имеющие какой-либо определенной формы. Другим ключевым моментом успеха подхода Хьюбеля и Визеля было то, что они искали не просто стимул, которые может вызвать ответ определенного нейрона, а наиболее эффективный для этого стимул. Исследование этой проблемы на разных этапах зрительной системы выявило много удивительных и неожиданных результатов. В ранних работах было показано, что рецептивные поля простых и сложных клеток в первичной зрительной коре составляют начальные этапы распознавания паттернов. Кроме того, анализ рецептивных полей четко выявил полезный и упрощающий принцип, что нейроны, расположенные вдоль радиальных трактов, идущих от поверхности коры в белое вещество, решают сходные задачи, формируя функциональные колонки. Эти результаты были окончательно подтверждены при помощи длинного электрода, вводимого далеко вглубь коры, одновременно с чем проводилась регистрация сигналов и идентификация свойств клеток, в зависимости от типа возбуждающего их стимула. После регистрации проводилась тщательная гистологическая реконструкция путей прохождения электрода.
Химический состав
По химическому составу бактерии не отличаются от клеток других организмов. Бактериальная клетка содержит 80% воды и 20% сухого остатка. Около 90% сухого остатка бактерии составляют высокомолекулярные соединения: нуклеиновые кислоты (10%), белки (40%), полисахариды (15%}, пептидогликон (10%) и липиды (15%); остальные 10% приходятся на мо ...
Кишечный сок
За сутки продуцируется около 2,5 л кишечного сока, принимающего участие в полостном гидролизе белков, углеводов, жиров.
В 12-перстной кишке продукция осуществляется за счет бруннеровых желез, расположенных в криптах, а в дистальной части этой кишки и па протяжении тощей и частично подвздошной - за счет либеркюновых желез, рН сока = 7,2 ...
Сравнение динамики развития фитопланктона за
2007-2008 года
В качестве сравниваемого материала служат 7 проб, взятых в 2007 году и 18 проб 2008 года, собранных в период с мая по октябрь.
В мае 2007 года наблюдалось более интенсивное развитие фитопланктона. По сравнению с численностью водорослей 1,95*109кл/м³ в 2007г в начале и середине месяца в 2008 году наблюдался уровень вегетации 0,819* ...