БиосинтезСтраница 1

После отделения арахидоновой кислоты от фосфолипида она выходит в цитозоль и в различных типах клеток превращается в разные эйкозаноиды. В клетках имеется 2 основных пути превращения арахидоновой кислоты: циклооксигеназный, приводящий к синтезу простагландинов, простациклинов и тромбоксанов, и липоксигеназный, заканчивающийся образованием лейкотриенов. (Рис.4)
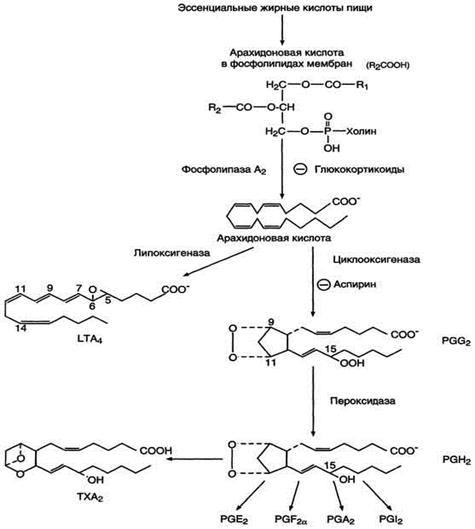
Рис.4 Синтез эйкозаноидов из арахидоновои кислоты.
Циклооксигеназный путь: синтез простагландинов и тромбоксанов
С20-три-, тетра - и пентаеновая жирные кислоты, служащие предшественниками простагландинов, входят в состав внутриклеточных фосфоглицеридов, из которых они высвобождаются под действием фосфолипазы А2. Активация фосфолипаз, ассоциированных с мембранами, происходит под действием многих факторов: гормонов, гистамина, цитокинов, механического воздействия. Этот процесс высвобождения предшественника, вероятно, играет роль регулирующей стадии в биосинтезе простагландинов, с помощью которой варьирует количество субстрата, подвергающегося последующему действию простагландинсинтазы. Последняя представляет собой связанный в мембране мультиферментный комплекс, содержащийся в микросомальной фракции тканевых гомогенатов. Необходимо отметить, что ингибиторное действие кортикостероидов на биосинтез простагландинов, связанное с торможением активности фосфолипазы А2, может служить объяснением антивоспалительного действия этих стероидных гормонов.
Фермент, катализирующий первый этап синтеза простагландинов, называется PG Н2 синтазой и имеет 2 каталитических центра. Один из них называют циклооксигеназой, другой - пероксидазой. Этот фермент представляет собой димер гликопротеинов, состоящий из идентичных полипептидных цепей. Фермент имеет гидрофобный домен, погружённый в липидный слой мембран ЭР, и каталитический домен, обращённый в полость ЭР. В активном центре циклооксигеназы находится тирозин (385), в активном центре пероксидазы - простетическая группа - гем. В организме имеются 2 типа циклооксигеназ (PGН2 синтаз). Циклооксигеназа 1 - конститутивный фермент, синтезирующийся с постоянной скоростью. Синтез циклооксигеназы 2 увеличивается при воспалении и индуцируется соответствующими медиаторами - цитокинами. Оба типа циклооксигеназ катализируют включение 4 атомов кислорода в арахидоновую кислоту и формирование пятичленного кольца. В результате образуется нестабильное гидропероксидпроизводное, называемое PG G2. Гидропероксид у 15-го атома углерода быстро восстанавливается до гидроксильной группы пероксидазой с образованием PG Н2. До образования PG Н2 путь синтеза разных типов простагландинов одинаков. Дальнейшие превращения PG Н2 специфичны для каждого типа клеток.
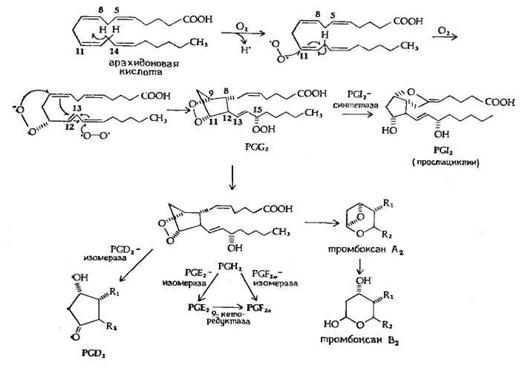
Рис.5. Предполагаемый механизм биосинтеза PGE2 и PGF2a из арахидоновой кислоты.
Биосинтез PGE2 и PGF2a, представленный на рис.5, начинается с арахидоновой кислоты и катализируется циклооксигеназой (М 70 000) жирных кислот, при этом образуется 15-оксиперокси-9-11-эндопероксид PGG2. Первая стадия в этой последовательности реакций приводит к потере атома водорода у С-13 с образованием свободного радикала. Эта стадия лимитирует скорость всего процесса. Затем происходят присоединение пероксидного радикала в положения 9 и 11 и образование 8,12 углерод-углеродной связи, которому предшествует изомеризация 11,12-двойной связи в 12,13-двойную связь и присоединение другого пероксидного радикала в положение 15, приводящее к изомеризации 12,13-двойной связи в 13,14-двойную связь. Все эти превращения рассматриваются как процесс одноэлектронного переноса, обозначенный на рисунке изогнутыми стрелками. Первый продукт этой последовательности реакций - это 15-оксиперокси-9,11-эндопероксид PGG2, существование которого было постулировано за несколько лет до его выделения и идентификации. PGG2 затем превращается в PGI2 или PGH2, вероятно, с помощью реакции пероксидазного типа. PGH2 является предшественником PGE2 и PGF2a в семенных пузырьках и других органах, а также тромбоксана А2 в тромбоцитах и легких.
Превращение арахидоновой кислоты в PGG2 блокируется ингибиторами циклооксигеназы жирных кислот. Наиболее известным среди этих ингибиторов являются аспирин (ацетилсалициловая кислота) и ряд других противовоспалительных лекарств (Рис.6). Торможение аспирином обусловлено переносом ацетильного остатка от молекулы аспирина к активному центру фермента с инактивацией последнего; такого рода механизм был предложен в качестве объяснения некоторых известных фармакологических эффектов аспирина.
Мочеполовая система
Мочеполовая система объединяет мочевые и половые органы. Они тесно связаны друг с другом по развитию, их выводные протоки соединяются у мужчин в общую трубку - мочеиспускательный канал, а у женщин открываются в общее пространство - преддверие влагалища. ...
Перечислите важнейшие различия между эукаритическими и прокариотическими
клетками
Характеристика
Прокариоты
Эукариоты
Размеры клеток
Диаметр в среднем 0,5-5 мкм
Диаметр обычно до 40 мкм Объем клетки в 103-105 раз больше, чем объем клетки эукариот.
Форма
Одноклеточные и нитчатые
Одноклеточные, нитчатые или многоклеточные.
Генетический материал
Кольцевая ДНК в цитоплазме, нет мате ...
Обеспечение организма витаминами
Витамины
(от лат. vita - жизнь) — вещества, которые не поставляют организму энергии, но необходимы в минимальных количествах для поддержания жизни. Они незаменимы и обычно поступают с растительной пищей или с продуктами животного происхождения, поскольку они не синтезируются клетками человеческого организма.
Важная роль в образовании ...