Глиоксилатный цикл. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение

Этот цикл в 1957 г. был впервые описан у бактерий и плесневых грибов Г. Л. Корнбергом и Г. А. Кребсом. Затем оказалось; что он активно функционирует в про- растающих семенах масличных растений и в других растительных объектах, где запасные жиры превращаются в сахара (глюконеогенез). Глиоксилатный цикл локализован не в митохондриях, как цикл Кребса, а в специализированных микротелах — глиоксисомах. В клетках животных этот цикл отсутствует.
В глиоксилатном цикле из ЩУК и ацетил-СоА синтезируется лимонная кислота, образуются цис-аконитовая и изолимонная (изоцитрат), как и в цикле Кребса . Затем изолимонная кислота под действием изоцитрат-лиазы распадается на глиоксиловую и янтарную кислоты. Глиоксилат с участием малатсинтазы взаимодействует со второй молекулой ацетил-СоА, в результате чего синтезируется яблочная кислота, которая окисляется до ЩУК.
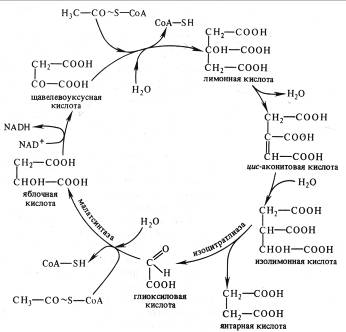
Рис. 6. Глиоксилатный цикл
Таким образом, в отличие от цикла Кребса в глиоксилатном цикле в каждом обороте участвует не одна, а две молекулы ацетил-СоА и этот активированный ацетил используется не для окисления, а для синтеза янтарной кислоты. Янтарная кислота выходит из глиоксисом, превращается в ЩУК и участвует, в глюконеогенезе (обращенном гликолизе) и других процессах биосинтеза. Глиоксилатный цикл позволяет утилизировать запасные жиры, при распаде которых образуются молекулы ацетил-СоА.
Регуляция ПФП и глиоксилатного цикла. Пентозофосфатный путь окисления регулируется концентрацией НАДФ + , так как содержит две НАДФ-зависимые дегидрогеназы (глюкозо-6- фосфатдегидрогеназу и 6-фосфоглюконатдегидрогеназу). Он регулируется также уровнем синтезов в клетке, потребляющих НАДФH (например, синтезом аминокислот и белков). Их высокий уровень приводит к увеличению содержания окисленного НАДФ + , что стимулирует ПФП.
В регуляции соотношения между ПФП и гликолизом принимает участие ряд интермедиатов: неорганический фосфат, 6-фосфоглюконовая кислота, эритрозо-4-фосфат. Недостаток неорганического фосфата подавляет гликолиз и активирует ПФП. 6-Фосфоглюконовая кислота служит ингибитором гликолитического фермента фосфофруктокиназы (глюкозофосфатизомеразы), что способствует функционированию ПФП. Эритрозо-4-фосфат, являясь субстратом транскетолазной и трансальдолазной реакций, может тормозить активность ферментов гликолиза и тем самым переключать превращения углеводов с гликолитического на пентозофосфатный путь.
Активность глиоксилатного цикла снижается при повышении концентрации оксалоацетата, который ингибирует сукцинатдегидрогеназу ЦТК. Другой ингибитор цикла — фосфоенолпируват подавляет активность изоцитратлиазы.
Влияние генов и среды на развитие признака
Примером влияния гена на общий метаболизм явл действие доминантного гена коротконогости у кур, который в гомозиготном состоянии летален, т.к.вызывает общие нарушения развития и гибель зародыша через 76 часов после начала инкубации. Примером влияния генов на отдельные биохимические реакции явл финилаланинтиразиновый обмен у человека. Исх ...
Неклассический этап
Неклассическое естествознание (конец XIX - середина XX в.в.) способствовало значительному расширению поля исследуемых объектов, открывая пути к освоению больших, сложных саморегулирующихся систем. Неклассический тип рациональности учитывает связи между знаниями об объекте и характером средств и операций деятельности, рассматривая объект ...
Почки и их типы. Происхождение и функции покоящихся, придаточных и спящих
почек
Почка представляет собой сильно укороченный побег с зачаточными листьями или репродуктивными органами. Почки могут быть вегетативные, имеющие зачатки побега и листьев; генеративные, несущие зачатки цветка или соцветия, смешанные. По местоположению различают почки верхушечные (на конце побега) и пазушные (в узлах между черешком листа и с ...