Первичный ответ и семейства рецепторов

Лиганд, связываясь с рецептором, должен индуцировать в нем конформационный переход, приводящий к функциональным изменениям в других частях молекулы. Природа такого перехода абсолютно неизвестна. Установлено лишь, что по крайней мере в IgE‑рецепторе с высоким сродством связывание лиганда индуцирует агрегацию рецептора в плоскости мембраны, однако нет никаких указаний на то, что это характерно для всех передающих сигнал рецепторов. Хотя структурные детали гормонов или нейромедиаторов остаются неизвестными, их важные функциональные особенности сейчас более или менее ясны. При связывании агониста происходят следующие три события.
1.Связывание агониста индуцирует открывание канала, образуемого рецептором. Примерами могут служить никотиновый ацетил-холииовый рецептор, рецептор 7‑амнномасляной кислоты и глициновый рецептор. Все это рецепторы нейромедиаторов, которые структурно объединены в малое суперсемейство. На основании данных о первичной структуре этих рецепторов / ионных каналов для каждого полипептида была построена модель с четырьмя трансмембранными сегментами, однако экспериментальные исследования топологии nAChR еще не закончены.
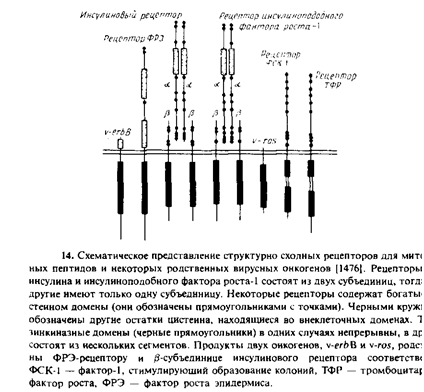
При связывании лиганда активируется тирозиновая протеин-киназа, представляющая собой цитоплазматический домен рецептора. Обычно сам рецептор и является мишенью, но данные о том, какие именно белки фосфорилируются и как они влияют на клетку, практически отсутствуют. Этот механизм используют рецепторы митогенных пептидных гормонов и факторов роста, причем многие из этих рецепторов структурно родственны между собой. Каждый из этих рецепторных полипептидов имеет по одному трансмембранному сегменту.
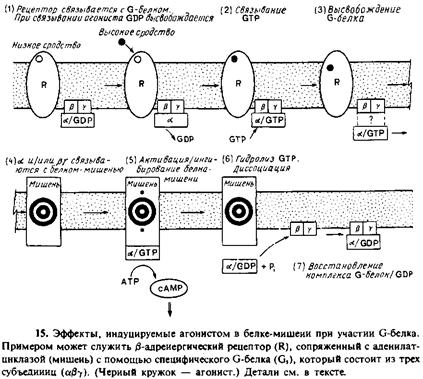
2.3. Рецептор образует комплекс с одной из групп мембраносвязанных GTP‑связывающих белков, называемых G‑белками. При связывании лиганда с рецептором в комплексе peuenTop/G‑белок происходит конформационное изменение, в результате чего облегчается обмен связанного GDP и GTP на G‑белке.
Последовательность происходящих событий изображена на рис. 9.15. G‑белок активируется на короткое время, будучи связанным с GTP, и в этом состоянии может отделиться от рецептора, причем одна или больше субъединиц G‑белка могут связаться с другими мембранными белками, обозначенными на рис. 9.15 словом «мишень», оказывая на них определенное воздействие. К этим мишеням относятся ионные каналы, аденилатциклаза, cGMP/фосфодиэстераза и фосфолипаза С.
Принадлежат также /З-адренергические рецепторы, мускариновые ацетилхолиновые рецепторы и опсины. Каждый из этих рецепторов имеет семь трансмембранных сегментов. На рис. 4.1 показана предполагаемая структура родопсина – наиболее полно охарактеризованного представителя этой группы.
Реакционная способность веществ, анализ и синтез. Реакционная способность веществ
РЕАКЦИОННАЯ СПОСОБНОСТЬ, характеристика относительной хим. активности молекул, атомов, ионов, радикалов. Для количественной оценки рассматривают реакционные серии, т.е. ряды однотипных реакций, проводимых в одинаковых условиях, например: (стандартная реакция)
где Х – общая группа атомов, которая претерпевает изменения в данной реакци ...
Разделение белков путем осаждения
Осаждение нуклеиновых кислот.
Обычно, когда говорят о высаливании, имеют в виду высаливание именно сульфатом аммония. Этому методу уже более 130 лет. Раньше он применялся и для фракционирования, сейчас, в основном, как дешёвый и удобный метод осаждения белков. Можно считать, что повезло, если при этом получается ещё и существенная очис ...
Метод определения
активности карбоксипептидазы М
Определение активности КП М проводится по выше изложенной схеме, но в качестве буфера используется 200 мМ трис-HCl, рН 7,4 [148]. ...