Гемагглютинин вируса гриппа – наиболее полно охарактеризован ная гликопротеиновая структура

Этот белок связан с вирусной мембраной с помощью короткого трансмембранного домена на С-конце. Он синтезируется как единая полипептидная цепь, но при созревании претерпевает протеолитическое расщепление с образованием двух полипептидов, HAi и НА2, связанных дисульфидной связью. Участок, отвечающий за слияние, локализован на N‑конце НА2. Он соответствует N‑концу G‑белка вируса везикулярного стоматита.
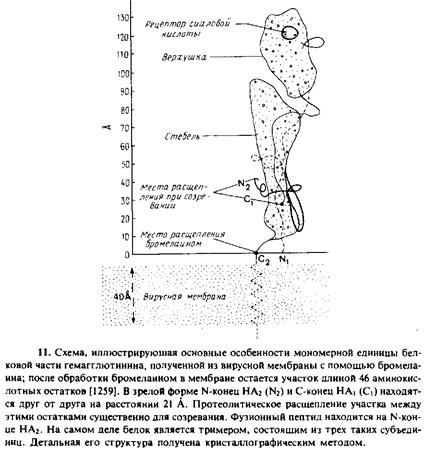
С помощью рентгеновской кристаллографии была определена трехмерная структура водорастворимого домена гемагглютинина. Этот домен получают с помощью расщепления бромелаином. Он представляет собой трнмер из – субъединиц, напоминает по форме стержень и выступает из мембраны на 135 А. Каждая субъединица имеет а-спиральный «стебель» с глобулярной «верхушкой», которая содержит рецепторыый участок для сиалогликосоединений.
Гидрофобный фузионный пептид спрятан между субъединицами тримера и находится на расстоянии 30 А от поверхности мембраны. Как известно, при низких рН третичная и четвертичная структуры белка необратимо изменяются. При рН 5,0 белок приобретает способность связывать липиды и детергенты и самоагретируется, что позволяет думать об экспонировании гидрофобного домена. По-видимому, это коррелирует с экспонированием фузионного пептида, который теперь может связаться с мембраной-мишенью, способствуя сближению обеих мембран и облегчая их слияние. Были выделены и получены in vitro мутанты с другими рН-оптнмумом и фузноинымн свойствами. Это подтверждает важность контактирования субъединиц при рН-зависимом конформационном изменении структуры белка и роль фузионного пептида. Гемагглютинин ацилирован жирными кислотами, которые, по-видимому, усиливают фузионную активность.
Суммируя все сказанное выше, можно сделать вывод, что, хотя физико-химические механизмы слияния мембран до конца не установлены, очевидно, что специфическое слияние мембран внутри клетки может осуществляться с помощью некоего белокзависимого процесса. Для этого должно произойти специфическое межмембранное взаимодействие, обеспечивающее прикрепление мембран друг к другу, и должен присутствовать мембранный белок, который при необходимости обеспечивает слияние. Для изучения свойств таких белков можно использовать белки, образующие шиловидные структуры вирусных частиц. Но могут применяться и другие модельные системы. Вероятно, семейство таких белков участвует в слиянии мембран как при экзоцитозе, так и при эндоцитозе.
Подсемейство нейротрофических факторов. Фактор роста нервов (ngf)
NGF был открыт и впервые исследован в 1951 году (Levi-Montalchini). Позднее фактор был идентифицирован в других тканях и у других видов, включая человека. В 1986 году Леви-Монтальчини получила совместно с С. Коэном Нобелевскую премию за работы по исследованию NGF.
Структура. Химические аспекты. NGF содержит 118 аминокислотных остатков, ...
Шишковидное тело
Шишковидное тело (corpus pineale), или эпофиз мозга — непарный орган массой 0,2 — 0,4 г, относится к эпиталамической области промежуточного мозга и располагается в неглубокой борозде, отделяющей друг от друга верхние холмики крыши среднего мозга. От переднего конца шишковидного тела к медиальной поверхности правого и левого зрительных б ...
Диморфизм и плеоморфизм
Для дрожжей, как и для других грибов, известны явления диморфизма и плеоморфизма. Мицелиально-дрожжевой диморфизм проявляется в том, что один вид может расти в двух формах – одноклеточной и мицелиальной. Это явление известно в микологии и связано с двумя типами роста – сферическим и апикальным. Есть дрожжи, которые образуют только однок ...