Сортировка компонентов комплекса
рецептор-лигандСтраница 1

В результате эндоцитоза и быстрого рециклирования плазматической мембраны происходит поглощение не только жидкостей. Одна из основных функций этого процесса состоит в облегчении поглощения внеклеточных белков. Большинство рецепторов, перечисленных в табл. 9.2, которые концентрируются в окаймленных ямках, связываются с такими макромолекулярными лигандами, как гликопротеины, липопротеины низкой плотности или иммуноглобулины. Кроме того, имеются рецепторы для факторов роста и таких гормонов, как ФРЭ или инсулин, когда в результате рециклирования плазматическая мембрана быстро освобождается от комплексов гормон–рецептор, заменяемых ново синтезированными рецепторами. Структурные детерминанты, которые опосредуют связывание этих рецепторов с окаймленными ямками, неизвестны. Установлена первичная структура нескольких таких рецепторов, при этом никакого сходства между ними не выявлено. Все они имеют единственный трансмембранный сегмент, но у одних N‑конец обращен в цитоплазму, а у других ориентирован в противоположном направлении. В нескольких случаях было показано, что цитоплазматический домен рецептора необходим для того, чтобы рецептор оставался в незамкнутых ямках, но об этом известно очень мало.
Судьбы рецептора и лнганда после их поглощения клеткой различаются. Исходя из этого можно выделить четыре основные группы систем лиганд–рецептор.
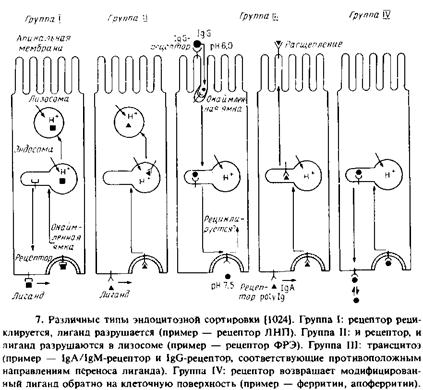
Группа I. Рецептор возвращается к поверхности клетки, лиганд направляется в лизосомы. Примерами служат ЛНП- и асиалоглико-протеиновый рецепторы.
Группа II. Как рецептор, так и лиганд направляются в лизосомы. Примерами являются ФРЭ- и инсулиновый рецепторы.
Группа III. Рецептор остается связанным с лигандом, а лиганд направляется в другой домен плазматической мембраны. Это явление называется трансцитозом и характерно для рецепторов иммуноглобулинов в поляризованных эпителиальных клетках. После такого клеточного «транзита» рецепторы для IgA и IgM подвергаются протеолитическому расщеплению, тогда как рецептор для IgG при некоторых условиях может использоваться вновь.
Группа IV. И рецептор, и лиганд возвращаются к одному и тому же домену плазматической мембраны. Наиболее полно охарактеризованным примером такого рода служит трансферриновая система, в которой после поглощения комплекса трансферрин–железо и удаления железа комплекс апотрансферрин–рецептор возвращается к клеточной поверхности.
Скорости всех этих процессов достаточно велики. Обычно время жизни рецепторов, возвращающихся к клеточной поверхности, составляет всего 3 – 6 мнн, а время полного оборота рецептора – около 15 – 25 мин. В равновесном состоянии на клеточной поверхности обнаруживается около 65 – 75% рецепторов, а остальная их часть находится на разных внутриклеточных мембранах. Полный анализ равновесной кинетики поглощения и процессинга может быть проделан с привлечением констант скоростей для каждого элементарного шага,
Одной из характерных особенностей механизма сортировки является его удивительная точность. Например, трансферриновый рецептор почти всегда находится в базолатеральной, но не в апикальной мембране поляризованных эпителиальных клеток. Вероятность его ошибочного включения в апикальную мембрану составляет лишь -0,1%, т. е. достаточно мала для сохранения полярности мембран этих клеток. В других системах, где по экспериментальным данным вероятность ошибки достигала 4%, для корректировки локализации рецепторов был необходим дополнительный этап сортировки.
Механизм сортировки должен быть весьма изощренным. По-видимому, различные комплексы рецептор–лиганд поглощаются вместе в одних и тех же частях окаймленных везикул, а сортировка внутри везикул происходит за несколько минут. Например, рецептор для асналогликопротеинов перемещается в латеральном направлении внутри эндосомной системы и концентрируется в трубочках и маленьких везикулярных компонентах, но не в крупных везикулах. Возможно, крупные эндосомные везикулы, практически лишенные этого рецептора, но содержащие гликопротеино-вый лиганд, сливаются затем с лизосомами. Однако трубочки, содержащие рецептор, возвращают его к клеточной поверхности. В этом случае сортировка несомненно происходит внутри эндосомного комплекса органелл.
Метод определения концентрации лактата в капиллярной крови пловцов
Величину лактата в крови определяли с помощью ферментного электрода с иммобилизованной лактатдегидрогеназой [26,34]. ...
Деление клетки. Митоз
Процесс деления у эукариот можно разделить на две стадии: митоз и цитокинез.
Митоз (от греч. «митос» – нить) – это образование из одного ядра двух дочерних ядер, морфологически и генетически эквивалентных друг другу. Цитокинез включает деление цитоплазматической части клетки с образованием дочерних клеток.
Биологическая роль митоза со ...
Активность
карбоксипетидазы Н и ФМСФ-ингибируемой карбоксипептидазы при введении
тестостерона в тканях самок мышей
Активность КП Н в гипофизе через 0,5 ч после введения тестостерона в дозе 3 мг на кг массы была выше на 61%, по сравнению с контрольной группой животных (рис. 3.2.1). Через 24 ч после введения тестостерона активность исследуемого фермента в гипофизе была на 16%, а в гипоталамусе и надпочечниках – на 45-50% ниже, чем у контрольных животн ...