Молекулы адгезии клетки и внеклеточного матрикса и
рост аксонаСтраница 1

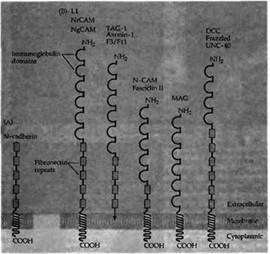
Молекулы клеточной адгезии управляют ростом аксона, обеспечивая наиболее благоприятное окружение для вытягивания конуса роста. Клеточные молекулы адгезии представляют собой трансмембранные или связанные с мембраной гликопротеины, характеризующиеся структурными мотивами своих внеклеточных частей, которые во многом гомологичны постоянным доменам иммуноглобулинов и фибронектина 3 типа (рис.2). Представителями надсемейства этих иммуноглобулинов (Ig) являются клеточные молекулы адгезии (N-CAM), молекулы адгезии нейроглии САМ (NgCAM), TAG-1, MAG, и DCC83). Эти молекулы обеспечивают адгезию клеток друг к другу посредством гетерофильных связей между различными надсемействами иммуноглобулинов (например, связь между NrCAM и TAG-1). Дополнительно присутствующей везде молекулой клеточной адгезии является молекула N-кадгерина (N-cadherin, рис. 2), которая обеспечивает кальций-зависимую адгезию между клетками.
В культуре клеток экспрессия N-CAM и N-кадгерина в клетках приводит к их агрегации, вытягиванию аксонов в сторону клеточных субстратов, но не субстратов внеклеточного матрикса, а также соединению отдельных растущих аксонов в пучки (фасцикулы, fascicles). Стимуляция роста аксонов молекулами клеточной адгезии не обеспечивается просто «липкостью» субстрата; она управляется активацией рецепторов, связанных с тиро1инкиназой, например рецептора к фактору роста фибробластов (fibroblast growth factor, FGF).
|
Рис. 2. Два класса молекул адгезии нервных клеток. (А) N-кадгерин способствует гомофильной кальций-зависимой адгезии. (В) Члены надсемейства иммуноглобулинов характеризуются многочисленными повторами связанных друг с другом дисульфидными мостиками циклов, которые гомологичны доменам, впервые описанным в постоянной части молекул иммуноглобулина. Многие из этих молекул клеточной адгезии также содержат несколько доменов, сходных с повторами в фибронектине III типа (показано в виде прямоугольников). У различных видов животных имеется большое количество гомологичных белков со сходными именами. |
|
Рецептор FGF включает внутриклеточный каскад, связанный с фосфорилированием тирозина, что приводит к удлинению аксона. Белковая тирозинфосфатаза, фермент, удаляющий остатки фосфата с тирозина, также участвует в регулировании этих сигналов. Молекулы адгезии внеклеточного матрикса,
включая ламинин, фибронектин, тенасцин (Л, цитотактин), а также перлецан, тоже являются благоприятным субстратом для роста нервных отростков85· 86). Эти гликопротеины с большой массой имеют две или более сходные субъединицы, удерживаемые вместе благодаря дисульфидным мостикам (рис. 2). Каждая субъединица характеризуется повторяющимися структурными мотивами. Белки внеклеточного матрикса взаимодействуют с клетками через семейство рецепторов, называемых интегринами. Было идентифицировано большое количество изоформ субъединиц интегрина. Каждая комбинация приводит к формированию рецепторов с различными свойствами. Интегрины обеспечивают структурные связи между белками внеклеточного матрикса и внутриклеточным актиновым цитоскелетом, регулируя форму клетки и ее миграцию. Кроме того, они активируют внутриклеточные сигнальные пути, которые управляют ростом клетки, пролиферацией и дифференцировкой.
Исследования при помощи специальных блокирующих антител показывают, что конусы роста редко используют только один субстрат для своего движения; несколько типов молекул адгезии клетки и внеклеточного матрикса могут обеспечивать рост нервных отростков у определенных типов нейронов. Например, для полного ингибирования роста аксонов в сторону шванновской клетки необходимо применять одновременно антитела к Ll/NgCAM, N-кадгерину и интегринам. Один тип антител сам по себе не может помешать росту аксона.
|
|
|
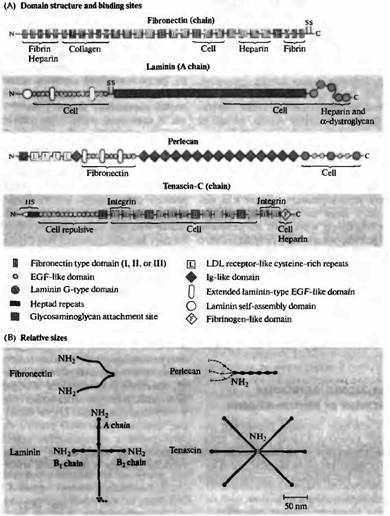
Рис. 3. Белки внеклеточного матрикса, опосредующие адгезию клеток к субстрату и отталкивание. (А) Схематическое представление доменной структуры цепей, состоящей из фибронектина, ламинина (А цепь), перцелана и тенасцина-С, а также участков, к которым присоединяются клетки, коллаген, гепарин, фибронектин, интегрин, фибрин и альфа-дистрогликан. S указывает на расположения остатков цистеина, которые образуют дисульфидные мостики между цепочками. Тенасцин-С содержит как связывающие, так и отталкивающие домены; основной его эффект, однако, отталкивание клеток и конусов роста. (В) Молекулы, изображенные в определенном масштабе. Все, кроме перцелана, являются олигопротеинами, цепи связаны друг с другом дисульфидными мостиками. Пунктирной линией показан гепарансульфат 90 и 170 нм длиной, связанный с N терминальным доменом перцелана. (Из Engel Л., 1992.) |
Критические периоды развития. Взаимодействие ядра и цитоплазмы в развитии
Взаимодействие ядра и цитоплазмы в развитии: цитоплазма играет важную роль в реализации наследственной инф-ции и формировании некоторых признаков организма. Основная часть цитоплазмы поступает в зиготу с яйцеклеткой. Определенные участки цитоплазмы яйцеклетки могут содержать факторы, определяющие судьбу тех или иных дифференцирующихся к ...
Диагностическое значение определения некоторых ферментов в крови
Определение активности ферментов все шире используется в диагностических целях в медицинской и ветеринарной практике. Этому способствует выяснение высокой информативности данного показателя для выявления патологического процесса на ранних стадиях заболевания, а также выпуск отечественных и зарубежных наборов необходимых реактивов, что з ...
Постановка реакции без использования фирменного набора(система SYBRgreen)
В качестве матрицы использовали плазмиду pcDNA3 cavmut (исходная концентрация 0,7 мг/мл), пару праймеров pc1 (исх.конц. 31мкМ) и pc2 (исх.конц. 12 мкМ), SYBRgreen и комплекс фермента (taq-полимераза) с антителами – так называемый Hotstart, 10-кратный сток буфера, растворы dNTP и MgCl2. Связывание полимеразы с антителами нужен для того, ...