Обновление фосфатидилинозитола и вторые
посредникиСтраница 1

Наиболее широкораспространенными мишенями G‑белков являются аденилатциклаза и фосфолипаза С, ответственная за гидролиз фосфатидилинозитола. Модуляция адени-латциклазы приводит к изменению внутриклеточной концентрации сАМР, который, как известно, служит вторым посредником, влияя на множество внутриклеточных процессов. Одним нз последствий увеличения содержания сАМР является, например, стимуляция сАМР-зависимой протеинкиназы, которая в свою очередь фосфорилирует специфические белковые субстраты. Клетки содержат также два типа Са2 + – зависимых протеннки-наз, активируемых соответственно Са2 +-кальмодулином и Са2 + вместе с диацилглицеролом и фосфатидилсерином. Активность обеих этих кии аз регулируется вторыми посредниками, образующимися при деградации фосфатидилинозитола, которая во многих клетках инициируется путем G‑белокзависимой активации специфической фосфолипазы С. Установление механизма обновления фосфатидилинозитола и физиологической роли продуктов его деградации явилось главным достижением в выяснении роли гормонов и нейромедиаторов в осуществлении клетками их функций.
На долю фосфатидилинозитола приходится лишь 2–8% всех фосфолипидов, содержащихся в клеточных мембранах эукариот. Структура его полярной головки представлена на рис. 9.16. Этот стереоизомер называют миоинозитолом, поскольку впервые он был выделен из мышц. Небольшая часть фосфатидилинозитола фосфорилирована по положению 4 или по положениям 4 и 5. От 1 до 10% фосфатидилинозитола, присутствующего в мембране, приходится на долю фосфатидилинози-толбисфосфата, обозначаемого как PIP2 или Р1Р2. Этот компонент является, вероятно, первой мишенью для фосфатидили-нозитолспецифичной фосфолипазы С, которая активируется во многих клетках G‑белком. Последующий гидролиз приводит к быстрому распаду фосфатидилинозитола в плазматической мембране и кратковременному возрастанию количества продуктов распада. Например, во время активации тромбоцитов за 90 с деградирует половина общего пула фосфатидилинозитола. Продукты распада действуют как вторые посредники и участвуют во многих клеточных процессах. Исследование этой системы еще не закончено, но общая ее схема уже построена и представлена на рис. 9.16. Последовательность событий такова.
1. Начальными продуктами гидролиза PIP2 являются диацил-глицерол и ииозитолтрисфосфат. Диацилглицерол связан с мембраной, а 1Рз является растворимым компонентом. Обычно жирные кислоты фосфатидилинозитола представлены стеариновой кислотой в положении 1 и арахидоновой кислотой в положении 2 глицерола. Из мозга быка были выделены две разные фосфатидилииозитолспецифичные фосфолипазы С, но данные об их активации G‑белком in vitro отсутствуют.
2. 1Рз служит вторым посредником, и его основной функцией, по-видимому, является мобилизация Са2 +, аккумулированного в эидоплазматическом ретикулуме. Возможно, этот компонент путем прямого связывания открывает Са2 + – специфичные каналы в эидоплазматическом ретикулуме, что приводит к увеличению концентрации Са2 + в цитоплазме в несколько раз. Обычно концентрация свободного Са2+ в цитоплазме составляет 0,1 мкМ.
3. Специфическая кии аза превращает некоторое количество 1Рз в тетрафосфорилированный продукт 1Р4. Образуются также другие, в том числе циклические, фосфоинозитиды, и некоторые из них тоже имеют физиологическое значение.
4. Одним из ферментов, регулируемых Са2 +, является фосфолипаза С, которая при низкой концентрации Са2+ использует в качестве субстрата преимущественно PIP2, но при более высокой концентрации Са 2 +, по крайней мере in vitro, использует нефосфорилированиый фосфатидилинозитол. Возможно, это облегчает непосредственный быстрый гидролиз основной части фосфатидилинозитола, но так ли это – неясно.
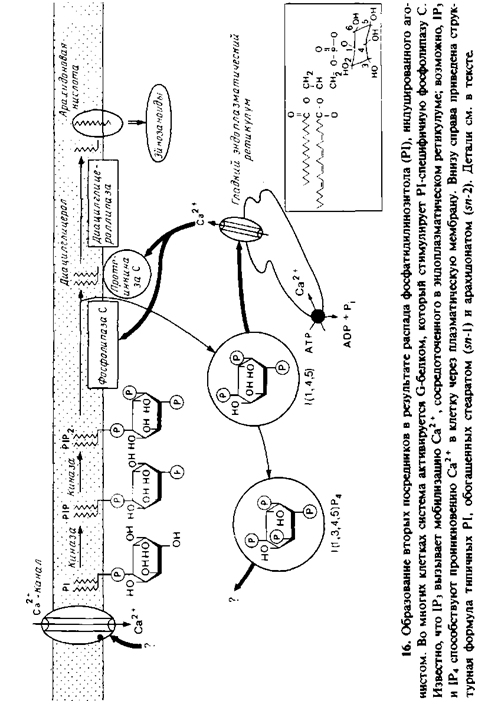
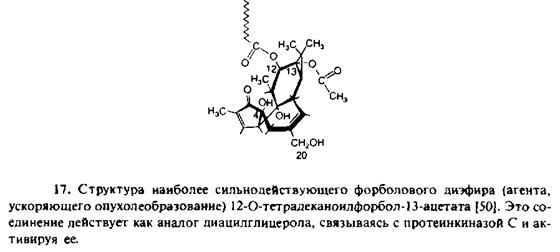
5. Наиболее важным ферментом, активируемым Са2 +, является протеинкиназа С. Этот фермент локализован преимущественно в цитозоле до момента появления там диацилглице-рола и Са2 +. Затем в зависимости от присутствия фосфатидилсери-на он связывается с плазматической мембраной и активируется. Эффекты, сходные с действием диацилглицерола, оказывают форболовые эфиры, и протеинкиназу С часто рассматривают как рецептор этих соединений. Одним из признаков участия протеинкиназы С и фосфоинозитидной системы в клеточном ответе на присутствие агониста является дублирование эффектов агониста путем добавления форболовых эфиров, которые непосредственно активируют протеинкиназу С.
Генетическая устойчивость к бесплодию
Бесплодие – нарушение воспроизводства потомства. Обусловлено многими факторами среды и генами. Во многих стадах гл причиной выбраковки – бесплодие. Степень проявления воспроизводительной способности варьирует от нормальной плодовитости до абсолютного бесплодия. Методами борьбы с бесплодием явл оценка генотипов производителей и интенсивн ...
Наследование, ограниченное полом. Наследование, контролируемое полом
Признаки, ограниченные полом: гены их наход-ся в аутосомах, т.е. есть у обоих полов, но проявляются только у одного пола. 1) Молочная продуктивность. 2) Яичная продуктивность. 3) Икра у рыб (у женского пола). 4) Яркое оперение (у самцов). К числу нежелательных, ограниченных полом признаков, относятся: 1) триторхизм, 2) аномалия спермиев ...
Принцип инвариантности
Смещение во времени и в пространстве не влияет на протекание физических процессов
. Здесь речь о переносах начала координат и начале отсчета времени. ...