Рецепторы, ответственные за хемотаксис Е. coli

К этому семейству белков относятся четыре рецептора. Их часто считают продуктами четырех генов – tsr, tar, tap и trg. Например, Wr‑белок связывается с аттрактантом серином, а также опосредует хемотаксис в ответ на репеллент лейцин. Наиболее полно изучен рецептор для аспартата, который связывается с аттрактантом аспартатом, а также с мальтозосвяззывающим белком. Все четыре рецептора содержат единственную полипептидную цепь. Данные об их первичной структуре позволяют предположить, что это трансмембранные белки, которые имеют по две пронизывающие мембрану а-спирали. Характер укладки рецептора для аспартата, представленный на рис. 9.12, согласуется с результатами экспериментов по слиянию генов, а также с генетическими и биохимическими данными. Биохимические свойства рецептора для аспартата свидетельствуют о том, что он, по-видимому, представляет собой тетрамер. Все четыре хемотаксических белковых рецептора имеют высококонсервативные С-концевые половины, составляющие цитоплазматические домены; N‑концевые части, составляющие периплазматические домены, гораздо менее консервативны.
Рецептор для аспартата выполняет три разные функции.
1. Связывание аспартата, за которое ответствен N‑концевой домен, обращенный в периплазму. Белки с укороченным цитоплазматическим доменом тем не менее связываются с аспартатом нормально.
2. Передача сигнала, в которой участвуют аминокислотные остатки трансмембранных спиралей; об этом свидетельствуют результаты замещения лизина в положении 19 на аланин в первой трансмембранной спирали. Полученный мутантный рецептор связывает аспартат, но не индуцирует ответной реакции, что, вероятно, обусловлено изменением конформации белковой молекулы. Показано также, что рецептор может связываться с аспар-татом и с мальтозосвязывающим белком одновременно, при этом ответы усиливают друг друга. Природа конформацнонного изменения в рецепторе, индуцируемого связыванием лиганда, неизвестна, но, по-видимому, это изменение затрагивает значительную часть полипептида. Механизм, с помощью которого рецептор влияет на «мотор», приводящий в движение жгутики, также неизвестен; установлено только, что в нем участвует С-концевой домен рецептора и он может быть связан с фосфорилнрованием одного из других белковых компонентов системы.
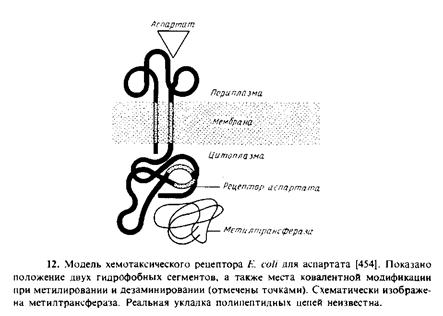
3. Адаптация, которая заключается в том, что система способна реагировать на увеличение концентрации лишь непродолжительное время; через несколько минут рецептор десенсибилизуется, т. е. адаптируется к новой концентрации аттрактанта. Но затем рецептор вновь приобретает способность реагировать на дальнейшие изменения концентрации аттрактанта. Частично это связано с метилированием и деметилированием рецептора по нескольким глутаминовым остаткам, расположенным в цитоплазматическом домене. Адаптация отсутствует у рецепторов с укороченным С-концом; эта мутантная форма реагирует на аспартат в течение всего времени, пока последний находится в среде.
В ходе многочисленных экспериментов была продемонстрирована кажущаяся независимость функциональных доменов, расположенных на N- и С-концах белка. Такие белки обладали хемотаксической активностью по отношению к серину. Подобные конструкции наблюдались также у семейства рецепторов пептидных гормонов животных клеток.
Из всего сказанного можно сделать следующие выводы.
1. Существует семейство трансмембранных рецепторов с родственными последовательностями.
2. Наличие у рецепторов разных функций предполагает, что у них имеются разные структурные и функциональные домены.
3. И высшие, и низшие организмы обладают системой передачи сигнала, в которую вовлечены другие цитоплазматические белки, вероятно каким-то образом модифицирующиеся благодаря конформационному изменению, которое претерпевает рецептор при связывании лиганда.
4. Система адаптируется к сигналу, т. е. может отвечать на него лишь непродолжительное время. Это свойство присуще многим системам клеточного ответа; его называют также десенсибилизацией.
Концепция самоорганизации объекта
Как и все остальные системы, заколка имеет свойство получать и выбрасывать потоки. Такое положение именуется как открытость систем.
На примере можно показать, что ряд потоков входящих в систему, при дальнейшем синтезировании, выбрасываются из неё, воздействуя на другие типы систем.
Подобное явление имеет ряд положительных и отрицатель ...
Показатели минерального обмена в плазме крови
Клиническое значение
Кальций Пониженное содержание кальция наблюдается при гипопаратиреозе, нарушении всасывания кальция, недостатке витамина D, при рахите, почечной недостаточности.
Повышенное содержание кальция отмечено при гиперпаратиреозе, гипертиреозе, злокачественных опухолях с метастазами в кости, при гипервитаминозе D. Опасным ...
Группы возбудителей инфекционных заболеваний
Регламентация условий работы с возбудителями инфекционных заболеваний произведена в соответствии со степенью опасности микроорганизмов для человека. По этому признаку выделено четы
ре
группы возбудителей.
Группа
I
:
возбудители особо опасных инфекций: чума, натуральная оспа, лихорадки Ласса, Эбола и др.
Группа
II
:
возбудители ...