Поверхность животной клетки

Прежде чем переходить к обсуждению структур различных мембраносвязанных рецепторов и механизмов индукции клеточного ответа, полезно рассмотреть строение поверхности животной клетки, поскольку большинство исследований в этой области выполнено на клетках животных. Плазматическую мембрану отличают от других клеточных мембран две особенности.
1. Относительно высокое содержание холестерола. Оценить долю холестерола клетки, находящуюся в плазматической мембране, довольно трудно, но ясно, что она весьма существенна. Высоким содержанием холестерола отличаются и другие мембранные структуры эндоцитозного пути. Еще один липид, содержащийся в значительных количествах в этих мембранах, – сфингомиелин.
2. Относительно высокое содержание гликопроизводных. Гликопроизводиые содержатся не только в плазматической мембране. Однако они присутствуют в ней в большом количестве. Все углеводные группы гликопротеинов плазматической мембраны находятся на наружной поверхности клетки, и зная места их прикрепления, можно определить аминокислотные остатки мембранных белков, обращенные наружу. Большинство белков плазматической мембраны гликозилированы. Полезно также помнить, что некоторые белки плазматической мембраны прикрепляются к ней с помощью фосфоинозитола и могут высвобождаться под действием фосфолипазы С, а другие белки присоединяются через ковалентно связанные жирные кислоты.
Физико-химические свойства холестерола в составе модельных мембран широко исследовались, однако функциональная роль холестерола в плазматической мембране до сих пор неизвестна. Далека от выяснения и биологическая функция углеводных частей гликолипидов и гликопротеинов.
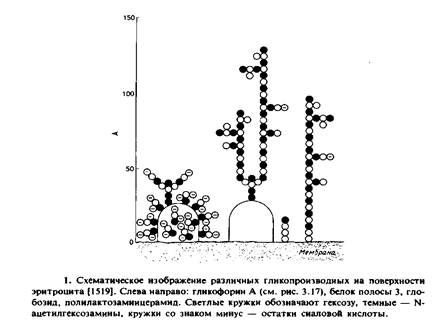
В качестве примера рассмотрим поверхность эритроцита. Два преобладающих в этой мембране белка – белок полосы 3 и гликофории А – являются достаточно хорошо охарактеризованными гликопротеинами. Около 75°7» всех моносахаридов клеточной поверхности входит в состав гликопротеинов, а остальные 25% содержатся в гликолипидах, которые состоят из простых глобозидов и длинноцепочечных полилактозаминцерамидов. Эти гликолипиды составляют лишь несколько молярных процентов от общего количества липидов. Основные углеводсодержащие компоненты мембраны эритроцитов схематически представлены на рис. 9.1; указана максимальная протяженность Углеводных цепей. Рис. 9.2 иллюстрирует поверхность эритроцита в истинном масштабе, дающем представление о расстоянии между различными гликопроизводными. На самом деле углеводные цепи не выступают над клеточной поверхностью так, как это изображено на рис. 9.1 и 9.2, однако эти структуры, вероятно, играют важную роль при взаимодействии клеток с их окружением.
Углеводсодержащие соединения придают клеточной поверхности ярко выраженный гидрофильный характер. При этом отрицательный заряд в значительной степени определяется остатками сиалоных кислот, которые составляют семейство производных нейраминовой кислоты. В некоторых случаях сиаловые кислоты маскируют специфические сайты узнавания на молекулах клеточной поверхности. Существенно, что гликопроизводные определяют свойства участка, выступающего над клеточной поверхностью на ~ 100 А. Кроме того, эти специфические компоненты обладают уникальными биологическими функциями.
Так, было показано, что углеводные компоненты гликолипидов и гликопротеинов изменяются при развитии и дифференцировке клетки и могут служить антигенными маркерами, ассоциированными с опухолями. Ганглиозиды, представляющие собой содержащие сиаловую кислоту гликосфинголипиды, служат местом связывания холерного и столбнячного токсинов. Что еще более интересно – ганглиозиды непосредственно участвуют в регуляции процессов клеточного роста и дифференцировки, а также, по-видимому, воздействуют на фосфорилирование рецептора фактора роста эпидермиса – возможно, путем прямого взаимодействия. При этом роль углеводного компонента гликопротеинов, и особенно рецепторных белков, не совсем ясна. Гликозилирование может играть модулирующую или регуляторную роль при функционировании некоторых рецепторов, необходимых для межклеточной адгезии.
Биология волнистого попугая. Характеристика отряда
Попугаи образуют хорошо выраженную, богатую видами группу птиц; характерным для них является строение клюва, который внешне похож на клюв хищных птиц, но более массивный и подвижный. Широкое и высокое у основания надклювье заканчивается заостренным крючком, нависающим над более коротким подклювьем. Подвижный клюв соединяется с лобными и ...
Органы выделительной системы
У низших многоклеточных (губки, кишечнополостные) обособленных органов выделения нет. У плоских и круглых червей появляются специальная выделительная система протонефридиального типа. У кольчатых червей - выделительная система метанефридиального типа, которая сохраняется в измененном виде у членистоногих.
Эволюция выделительной системы ...
История зоопарка
Московский зоологический сад был торжественно открыт 31 января 1864 года. Инициатором создания зоопарка было Русское Императорское Общество Акклиматизации растений и животных. Царское правительство выделило под строительство зоопарка землю на окраине Москвы вокруг Пресненских прудов, а также 10 000 рублей золотом. Значительная часть сре ...