Навигация конусов роста в спинном мозгеСтраница 1

Хороший пример разнообразия механизмов и молекул, которые используют конусы роста для навигации к своим клеткам-мишеням, представляют аксоны комиссуральных интернейронов спинного мозга позвоночных В раннем периоде развития комиссуралъные интернейроны, которые лежат в дорзальной части спинного мозга, «выпускают» аксоны, которые начинают расти в вентральном направлении, пересекая среднюю линию, и затем растут вдоль спинного мозга по направлению к своим синаптическим мишеням (рис. 6) .
Аксоны комиссуральных нейронов первоначально привлекаются к вентральной средней линии белком нетрин-1, растворимым хемоаттрактантом, образуемым специальными клетками базальной пластинки, лежащими вдоль средней линии спинного мозга (рис. 23.24А)111). Нетрин-1 взаимодействует с рецептором, экспрессируемым комиссуральными нейронами, называемым DCC (который уже упоминался ранее из-за своего взаимодействия с молекулами клеточной адгезии. Существование растворимого фактора, образующегося в базальной пластинке, который способен привлекать (attract) аксоны комиссуральных нейронов, было впервые показано при культивировании кусочков дорзальной части спинного мозга изолированно, либо вместе с кусочками базальной пластинки (рис. 7). Аксоны комиссуральных нейронов растут четко по направлению к базальной пластинке, даже если эксплантаты находятся друг от друга на расстоянии нескольких сотен микрон. Эта дистанция слишком велика для того, чтобы ее прошел филоподий конуса роста и требует растворимого фактора. Такой фактор, нетрин-1, входящий в семейство секреторных белков, характеризуется сходством доменов с N-концевыми доменами 7 цепи ламинина-1 (рис. 7А). Гомологи нетрина также играют роль в росте аксонов у дрозофилы и С. elegans.
Далее аксоны комиссурального нейрона пересекают вентральную среднюю линию, что они делают только однажды, таким образом оставаясь на контралатеральной стороне (рис. 7В, С). Это пересечение облегчается взаимодействиями между двумя адгезивными молекулами клеточной поверхности: TAG-1, которая экспрессируется на поверхности аксона спаечного нейрона, и NrCAM, экспрессируемого на клетках базальной пластинки. После пересечения средней линии экспрессия TAG-1 в аксонах ингибируется сигналами от клеток базальной пластинки, под действием которых в аксонах начинает синтезироваться протеин, называемый robo. Robo представляет собой рецептор для другого белка, называемого slit, который высвобождается клетками базальной пластинки. Взаимодействие между slit и robo отталкивает конусы роста комиссуральных интернейронов. Потеря аттракции к TAG-1—NrCAM контакту и приобретение коротко-дистантного отталкивающего взаимодействия slit-robo предупреждают возможное повторное пересечение аксоном средней линии.
|
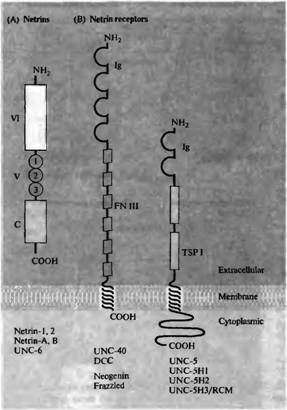
Рис. 8. Роль нетрина и его рецепторов в аттракции и отталкивании на большом расстоянии. (А) N-концевая аминокислотная часть секреторного белка нетрина состоит из доменов VI и V, которые гомологичны доменам аминокислотного конца 7 цепи ламинина. Домен V содержит три ECG-повтора. С-концевой домен этой цепи не имеет гомологии с ламинином. (В) Рецепторы нетрина имеют внеклеточный домен, однократно пересекающий мембрану, и внутриклеточный домен. Внеклеточный домен семейства DCC рецепторов нетрина имеет четыре иммуноглобулиновых домена (Ig) и шесть повторов фибронектина III (FN III). DCC и их гомологи также могут играть роль молекул адгезии. Семейство рецепторов к нетрину UNC-5 имеет два иммуноглобулиновых домена и два домена, гомологичных тромбоспондину 1-го типа (TSP I), которые расположены внеклеточно, а также длинную цитоплазматическую последовательность. (С) Микрофотографии кусочков дорзального спинного мозга эмбрионов крысы (сверху на каждой из панелей), культивированных вместе с кусочками ткани базальнои пластинки (слева), вместе с CDS клетками, секрета рующи ми рекомбинантный нетрин 1 (в центре), и с контрольными COS клетками (справа). Нижняя пластинка и нетрин 1 оба способны вызывать значительный и направленный рост пучков комиссуральных аксонов из дорзаль ной области спинного мозга. |
|
Регуляция транскрипции у про- и эукариот
Экспрессия генов как у прокариот, так и у эукарит регулируется при помощи целого ряда механизмов. Некоторые из механизмов такого рода, действующие в бактериальных системах, изучены довольно хорошо, но о том, как действуют регуляторные механизмы в клетках эукариот известно немного.
Осуществляя контроль за тем, каким генам экспрессироват ...
Общая характеристика
Самая древняя и многочисленная группа хордовых животных. Около 22 тыс. видов, заселивших моря, океаны и пресные воды. В течении длительного процесса эволюции рыбы совершенствовали свои гидродинамические приспособления и в результате приобрели обтекаемую торпедообразную форму тела, сплющенную с боков или в спинно-брюшном направлениях. Пе ...
Цвет колоний
При просмотре посевов также обращают внимание на цвет колоний. Чаще они бесцветные, белые, голубоватые, жёлтые или бежевые; реже — красные, фиолетовые, зелёные или чёрные. Иногда колонии ирризируют, то есть переливаются всеми цветами радуги [от греч. iris, радуга]. Окрашивание возникает в результате способности бактерий к пигментообразо ...