Путь углерода в фотосинтезе. Цикл Кальвина

Включение С02 в конструктивный метаболизм у фототрофов осуществляется в последовательности реакций. Она называется циклом автотрофной фиксации углекислоты, или циклом Кальвина, который сходен с пентозофосфатным циклом катаболизма сахаров у хемоорганогетеротрофов. Приведем суммарное уравнение этих реакций:
6С02 + 18 АТР + 12 NADPH + 12Н+ -> С6Н1206 + 18ADP + 12NADP+.
Для синтеза одной молекулы глюкозы здесь требуется 6 «оборотов» цикла. В цикле Кальвина, по сравнению с пентозо-фосфатным циклом, для регенерации акцептора С02 необходимы две дополнительные реакции:
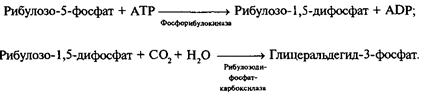
Дальнейшая последовательность реакций представляет собой «обращение» гликолиза, где в качестве восстановителя используется NADPH, что приводит к образованию глюкозы.
Аналогичный цикл фиксации С02 функционирует у хемоли-тоавтотрофных бактерий.
У растений начальные ферменты цикла Кальвина локализованы в хлоропластах. В клетках фототрофных бактерий рибулозоди-фосфаткарбоксилаза найдена в карбоксисомах, хотя не исключено, что там она пребывает в латентном состоянии.
В процессе функционирования цикл Кальвина подвергается строгой метаболической регуляции. Особенно тонко регулируется активность фосфорибулокиназы, в частности, чувствительной к энергетическому заряду клетки и к степени восстановленности NAD. Цикл работает эффективно только в условиях нормального снабжения энергией и восстановителями.
Бесхлорофильный фотосинтез
Особый тип фотосинтеза – без участия хлорофильных пигментов – обнаружен у экстремально галофильных архебактерий. Это единственный тип фотосинтеза, не включающий электрон-транспортную цепь. Клетки галобактерий содержат особый белок – бактериородопсин, включающий каротиноид – ретиналь. Совместно с фосфолипидами бактериородопсин формирует так называемые фиолетовые мембраны, которые в виде «бляшек» покрывают до 50% поверхности клеток. Под действием света бактериородопсин транслоцирует протоны из цитоплазмы в окружающую среду, создавая протонный ТЭП, который посредством Н+-АТРсин-тетазы преобразуется в АТР.
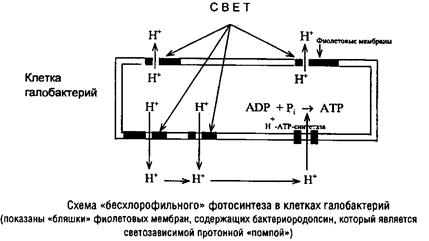
Система автотрофной фиксации С02 у галобактерий не обнаружена, по-видимому, они существуют как фотоорганогетеротрофы.
Органическая химия
ОРГАНИЧЕСКАЯ ХИМИЯ, наука, изучающая соединения углерода с другими элементами (органические соединения), а также законы их превращений. Название "органическая химия
" возникло на ранней стадии развития науки, когда предмет изучения ограничивался соединениями углерода растительного и животного происхождения. Не все соединения у ...
Биологическая характеристика европейской ряпушки в связи со средой обитания
и образом жизни. Общая биология вида
Тип: Хордовые (Chordata)
Под тип: Черепные (Craniata)
Над класс: Челюсноротые (Gnathostomata)
Класс: Рыбы (Pisces)
Над отряд: Костистые рыбы (Teleostei)
Отряд: Лососеобразные (Salmoniformes)
Под отряд: Лососевидные (Salmonоidei)
Семейство: Сиговые (Coregonidae)
Род: Сиги (Coregonus)
Вид: Европейская ряпушка (Coregonus albula L) ...
Показатели жирового обмена в сыворотке крови
Клиническое значение.
Определение в крови общих липидов и их отдельных фракций рекомендуется проводить в крови, взятой через 12 час. после еды. В противном случае повышенное содержание липидов (гиперлипидемия) не имеет диагностического значения. При учете сказанного выше гиперлипидемия может указывать на поражение печени и почек, в осо ...